Home > D. Systemic pathology > Genetic and developmental anomalies > Genetic metabolic diseases > iron metabolism
iron metabolism
Wednesday 25 June 2003
Iron is fastidiously conserved by living cells since it is required for aerobic respiration; however, in excess the metal is toxic.
In mammals no effective excretory pathway exists and iron homeostasis is uniquely regulated by the intestinal mucosa, which controls the uptake and transfer of iron from the diet to meet the needs of growth and to recover obligatory losses.
Iron absorption
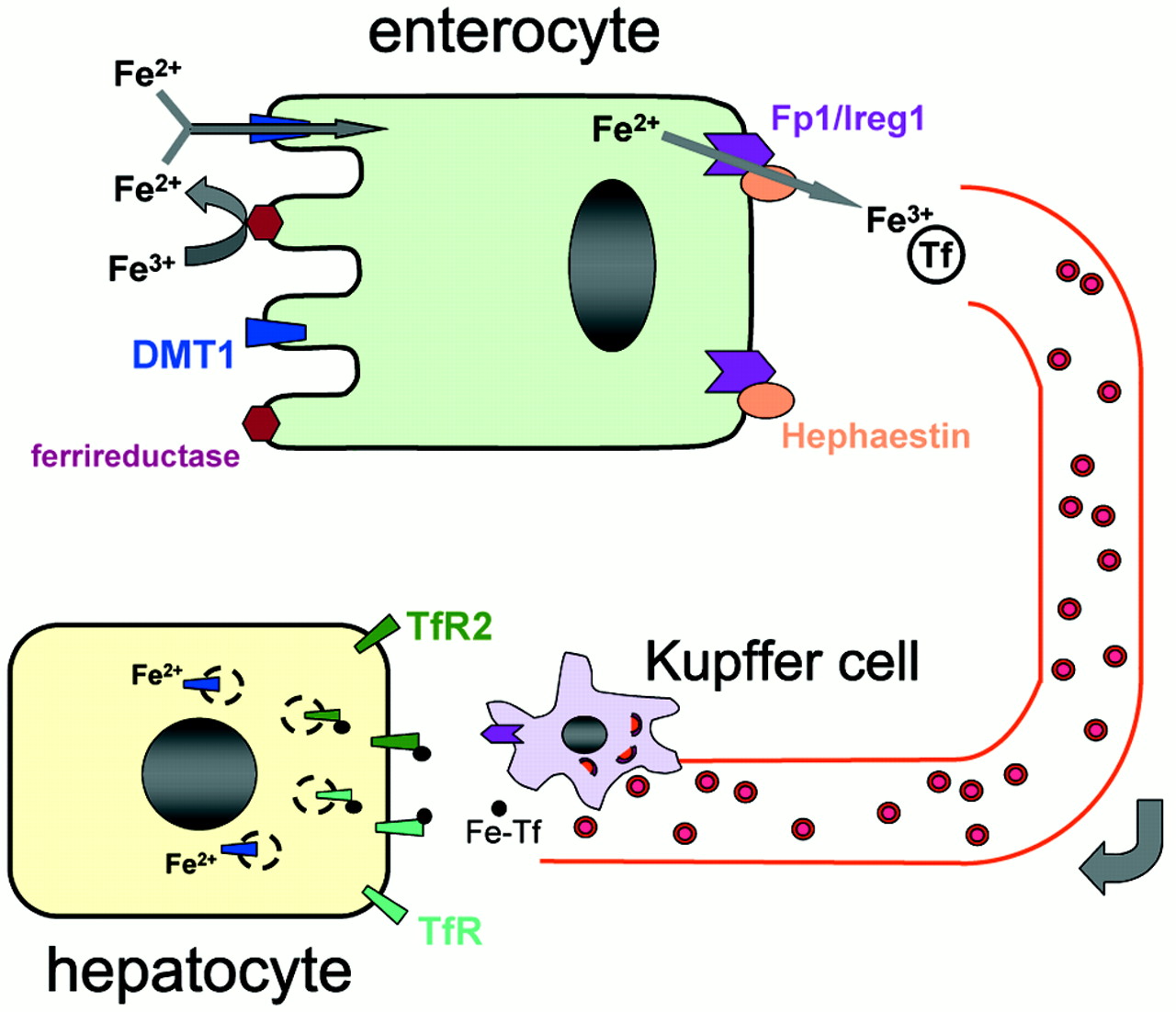
Iron can be absorbed from the intestinal lumen either as ferrous iron or as heme. The absorption of heme is not well understood, but may well be a yet-to-be elucidated active process.
The heme and non-heme iron pathways presumably converge after heme oxygenase in the mucosal cell opens the porphyrin ring and releases the iron. Ferrous iron probably enters the cell luminal surface through the divalent metal transporter 1 (DMT1).
Ferric iron must be reduced before traversing the membrane through DMT1, and this is accomplished by an intestinal cytochrome that functions as a ferrireductase and that is designated DCYTB.
It has been proposed that ferroportin is also present on the microvilli of the intestinal mucosal cell and that it may play a role in iron absorption.
A system consisting of mobilferrin and an integrin that may serve as a transport system for luminal ferric iron has also been described.
Some of the iron gaining entrance to the cell may be incorporated into ferritin. At one time it was believed that this ferritin iron caused a “mucosal block,” a concept that was flawed from the beginning.
No such block exists, although the mucosa does respond appropriately to iron lack and iron excess.
Export of ferrous iron from the cell at the basolateral membrane occurs through ferroportin (IREG1, MTP1). During this process iron is reoxidized to the ferric form through the action of hephaestin, a copper-containing ceruloplasmin analog.
Exactly how the iron exported from the cell reaches its binding site on serum transferrin is unknown. The transport of iron from cells by ferroportin is an important limiting step—one that seems to be compromised in almost all forms of hereditary hemochromatosis known so far.
In most types of hemochromatosis, the plasma transferrin saturation is greatly increased, and not only immediately after ingestion of iron. It therefore appears that the abnormality of iron transport in hemochromatosis is generally not limited to the mucosal cells but includes other tissue cells.
Under normal circumstances, an accurate mechanism regulates total body iron stores. When the amount of iron in the body is insufficient for its needs, absorption is upregulated. When the iron content is excessive, absorption is downregulated. The latter mechanism is also invoked by infection; microorganisms require iron for growth, and depriving them of this vital element is one defense mechanism against infection.
Iron homeostasis
Plasma levels of iron are closely regulated to ensure a daily supply of approximately 20 mg to the bone marrow for incorporation into hemoglobin in erythroid precursors and mature red cells.
Most of the iron found in the plasma derives from the continuous breakdown of hemoglobin in senescent red cells by reticuloendothelial macrophages.
Approximately 1 to 2 mg per day is also taken up by duodenal enterocytes and transferred to the plasma compartment or, depending on body needs, stored in the enterocytes as ferritin.
These stores are eliminated when enterocytes are sloughed at the end of their life cycles; apart from menstrual blood loss, this is the only significant means by which excess body iron is excreted.
Iron recycled by macrophages (as well as that absorbed from the gut) is loaded onto serum transferrin and delivered primarily to the bone marrow for reincorporation into new red-cell precursors.
The remaining body iron (approximately 1000 mg) is stored, primarily in hepatocytes.
Iron metabolism anomalies (iron-linked diseases)
![]() primary iron overload disorder (primary hemochromatoses)
primary iron overload disorder (primary hemochromatoses)
- classic hemochromatosis (hemochromatosis type 1 or HFE1) (HFE mutations)
- juvenile hemochromatosis (hemochromatosis type 2 or HFE2)
- hemochromatosis type 3 (HFE3)
- hemochromatosis type 4 (HFE4)
![]() secondary iron overload disorders
secondary iron overload disorders
The identification of HFE, the principal determinant of adult haemochromatosis (HFE1) (MIM.235200) and TfR2, recently implicated in a rarer form of the inherited disorder (HFE3) (MIM 604250), and the promise of candidate genes for juvenile haemochromatosis (HFE2; OMIM 602390) and neonatal haemochromatosis (MIM.231100) provide the foundation for important studies into the control mechanism of iron balance in humans.
The rare conditions atransferrinaemia (MIM.209300) and acaeruloplasminaemia (MIM.604290), each associated with tissue iron overload, have already implicated the iron transport ligand transferrin and the copper transporter caeruloplasmin in the control of iron homeostasis.
Animal models
Gene mapping studies in animal mutants with anaemia due to defects in the uptake or tissue transfer of iron have yielded novel proteins involved in iron transport: ![]() DMT1 (brush border transporter of ferrous iron) in the mk/mk mouse, hephaestin (basolateral multi-copper ferroxidase) in the sex-linked anaemic mouse (sla)
DMT1 (brush border transporter of ferrous iron) in the mk/mk mouse, hephaestin (basolateral multi-copper ferroxidase) in the sex-linked anaemic mouse (sla) ![]() ferroportin1 (basolateral iron exporter) in zebrafish weh mutants.
ferroportin1 (basolateral iron exporter) in zebrafish weh mutants.
See also
![]() iron acquisition
iron acquisition![]() iron storage
iron storage![]() iron transport
iron transport![]() intracellular iron
intracellular iron
References
![]() Lee DW, Andersen JK. Role of HIF-1 in iron regulation: potential therapeutic strategy for neurodegenerative disorders. Curr Mol Med. 2006 Dec;6(8):883-93. PMID: 17168739
Lee DW, Andersen JK. Role of HIF-1 in iron regulation: potential therapeutic strategy for neurodegenerative disorders. Curr Mol Med. 2006 Dec;6(8):883-93. PMID: 17168739
![]() De Domenico I, McVey Ward D, Kaplan J. Regulation of iron acquisition and storage: consequences for iron-linked disorders. Nat Rev Mol Cell Biol. 2008 Jan;9(1):72-81. PMID: 17987043
De Domenico I, McVey Ward D, Kaplan J. Regulation of iron acquisition and storage: consequences for iron-linked disorders. Nat Rev Mol Cell Biol. 2008 Jan;9(1):72-81. PMID: 17987043
![]() Ganz T. Molecular control of iron transport. J Am Soc Nephrol. 2007 Feb;18(2):394-400. PMID: 17229910
Ganz T. Molecular control of iron transport. J Am Soc Nephrol. 2007 Feb;18(2):394-400. PMID: 17229910
![]() Zecca L, Youdim MB, Riederer P, Connor JR, Crichton RR. Iron, brain ageing and neurodegenerative disorders. Nat Rev Neurosci. 2004 Nov;5(11):863-73. PMID: 15496864
Zecca L, Youdim MB, Riederer P, Connor JR, Crichton RR. Iron, brain ageing and neurodegenerative disorders. Nat Rev Neurosci. 2004 Nov;5(11):863-73. PMID: 15496864
![]() Roy CN, Andrews NC. Recent advances in disorders of iron metabolism: mutations, mechanisms and modifiers. Hum Mol Genet. 2001 Oct 1;10(20):2181-6. PMID: 11673399
Roy CN, Andrews NC. Recent advances in disorders of iron metabolism: mutations, mechanisms and modifiers. Hum Mol Genet. 2001 Oct 1;10(20):2181-6. PMID: 11673399
![]() Qian ZM, Shen X. Brain iron transport and neurodegeneration. Trends Mol Med. 2001 Mar;7(3):103-8. PMID: 11286780
Qian ZM, Shen X. Brain iron transport and neurodegeneration. Trends Mol Med. 2001 Mar;7(3):103-8. PMID: 11286780
![]() Andrews NC. Iron homeostasis: insights from genetics and animal models. Nat Rev Genet. 2000 Dec;1(3):208-17. PMID: 11252750
Andrews NC. Iron homeostasis: insights from genetics and animal models. Nat Rev Genet. 2000 Dec;1(3):208-17. PMID: 11252750
![]() Griffiths W, Cox T. Haemochromatosis: novel gene discovery and the molecular pathophysiology of iron metabolism. Hum Mol Genet. 2000 Oct;9(16):2377-82. PMID: 11005792
Griffiths W, Cox T. Haemochromatosis: novel gene discovery and the molecular pathophysiology of iron metabolism. Hum Mol Genet. 2000 Oct;9(16):2377-82. PMID: 11005792
![]() Bottomley SS. Secondary iron overload disorders. Semin Hematol. 1998 Jan;35(1):77-86. PMID: 9460811
Bottomley SS. Secondary iron overload disorders. Semin Hematol. 1998 Jan;35(1):77-86. PMID: 9460811
{kind=link}